
First, a little background. Centrosomes are one of the important organelles in the cell. During cell division centrosomes divide and each centrosome moves to an opposite end off the cell. Microtubules extend out and connect to the chromosomes and as the cell divides, the microtubles (called a spindle at this point) pull the (duplicated) chromsomes into their half of the cell. The microtubules themselves are polarized - that is they have a positive and negative end. The positive end is the one that attaches to the chromosomes (and kinetochore).
Centrosome and Centrioles

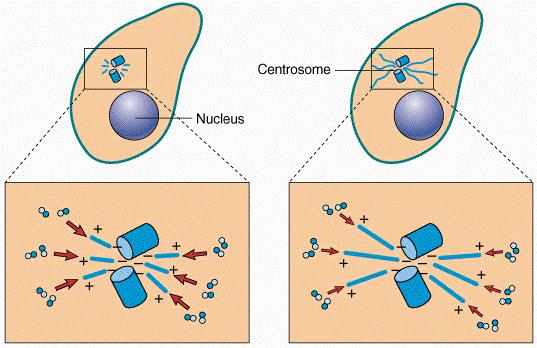
Centrosomes also play a major role in microtubule origination (called nucleation), elongation (called polymerization) and degradation (depolymerization).

Situated within each centrosome are a pair of centrioles. Surrounding each centriole pair in the cytoplasm is, what appears to be, a network of small fibers. The centrioles are arranged at right angles to each other.
Centrioles
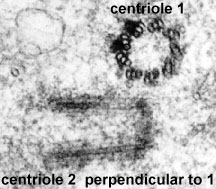
The centrioles themselves are made of nine groups of three microtubules, which form the walls of the centriole. Each triplet is linked it's neighbors at various spots along the length of the microtubules. Each triplate is also tilted inward - like the blade of a turbine. One of the centrioles has another feature - called the subdistal appendages - on each triplet. The subdistal appendages serve to anchor the microtublues forming the spindle. Surrounding each centriole is a region containing pericentrioler material which serves to nucleate the microtubules. The paired centrioles are also linked by interconnecting fibers.
Centrioles

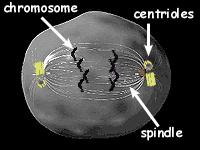
Below is a picture of a cell in the process of dividing.
Centrioles, Spindles and Chromosomes
With that in mind let's examine Well's argument concering centrioles. A subscription is required to view the Rivista Di Biologiaarticle. However, you can find one version of it Using Intelligent Design Theory to Guide Scientific Research. Wells' claims that because the microtubule triplets are tilted inwards just like turbine paddles they must actually be turbines.
In the electron microscope, centrioles look like tiny turbines. Using TOPS
as my guide, I concluded that if centrioles look like turbines they might actually
be turbines. I then used reverse engineering to formulate a testable, quantitative
hypothesis linking centrioles, polar ejection forces, and cancer.
He argues that the distal appendages form a flange which holds the centriole in its (sub distal appendage) bearing. He points out that when deprived of calcium the subdistal appendages dissasociate from the centriole (thus interfering with cell division). He also claims that one of the centrioles is a "helical pump" while the other centriole "would not rotate on its own axis; instead, it would swing bodily around the long axis of the mother centriole." Wells then compares this to a vortexer (used to centrifuge different materials) and claims that create a "polar ejection force" that "
subjects chromosomes to unusual stresses that cause breaks and translocations.
Even more serious than the presence of extra centrioles would be a failure of the
control mechanisms that normally shut down centriolar turbines at the beginning
of anaphase, since centriole pairs would then continue to accelerate and generate
polar ejection forces far greater than normal." He then notes a link between calcium deficiency (see above) and cancer and claims he has found one possible cause. Is this true? Before I answer that question let me point out two more statements Wells made in the article.
TOPS (Theory of Organismal Problem Solving)then explicitly rejects several implications of Darwinian evolution.
These include: (1a) The implication that living things are best understood from
the bottom up, in terms of their molecular constituents. (1b) The implications
that DNA mutations are the raw materials of macroevolution, that embryo
development is controlled by a genetic program, that cancer is a genetic disease,
etc.
And:
Finally, TOPS assumes as a working hypothesis that various implications
of ID are true. These include: (2a) The implication that living things are best
understood from the top down, as irreducibly complex organic wholes. (2b) The
implications that DNA mutations do not lead to macroevolution, that the
developmental program of an embryo is not reducible to its DNA, that cancer
originates in higher structural features of the cell rather than in its DNA, etc.
I have bolded the important parts. Let's examine the links between DNA and Cancer (which Wells specifically rejects) first.
I quickly learned from reviewing the recent scientific literature that cancer is not correlated with any consistent pattern of DNA mutations, but it is
correlated with abnormalities at the chromosomal level -- a phenomenon called
"chromosomal instability" (Lengauer et al., 1998). Chromosomal instability, in
turn, is correlated with centrosome abnormalities -- particularly the presence of
extra or enlarged centrosomes.
This is semi-true as far as it goes. Chronic Myelogenous Leukemia, is caused by a translocation between the long arms of the 9th and 22nd chromosome. On the other hand, it completely ignores the work on Oncogenes and Proto-Oncogenes. It also ignores work done in the area of retroviruses as well as in karyotyping of tumor cells. In several cases all three approaches converged (i.e identified) on the same DNA abnormalities. As another example, consider retinoblastoma. Some people with a hereditary form of retinoblastoma have a deletion of a specific band on chromosome 13, this same deletion is also encountered in tumor cells from people with a non-hereditary form. This gene has been called Rb. In patients with the hereditary form of the cancer Normal cells are defective in one of two copies of the gene while the cancerous cells are defective in both copies of the gene. In the nonhereditary form non-cancerous cells have two normal copies of the Rb gene while cancerous cells have two defectivve copies of the Rb gene. Further research on the issue has implicated the Rb gene in a number of different cancers (mainly cancers of the lung, breast, and bladder). One of the products of Rb acts as a break on the progress of cell division. This is regulated by the phosphorylation of the Rb protein. Unphosphorylated Rb binds to rene regulatory proteins and prevents them from promoting DNA replication (and hence cell division and proliffereation). There are several other examples that could be given, but at this point that would be redundant. Wells is completely wrong on the connection between DNA and cancer.
Cancer has several different causes though, so might night Wells' centriole as a turbine/polar ejection theory be correct? The answer is no. Abnormal Spindles:
Before a normal cell divides, its chromosomes are duplicated and then pulled apart by a structure called a spindle, so that the two daughter cells each will have the same number of chromosomes.
At the end of a normal spindle is the spindle pole, also called the centrosome, which pulls the chromosomes outward. Cancer cells often have extra centrosomes. When a cell has more than two centrosomes, sometimes--but not always--the spindles will have more than one pole and cell division won't work correctly, leading to the swapping of genetic material, uncontrolled cell division, and the formation of tumors.
Why this doesn't always happen when there are too many centrosomes was the focus of the Pitt researchers' investigation. They found that as long as the extra centrosomes "cluster" together, the spindles will form normally, with two ends, and the cells will divide normally. "No one else appreciated that that was required, or what the mechanism was that separated them," said Saunders.
But when the extra centrosomes don't cluster together, the spindles don't form normally, and cell division can become unstable.
Investigating the mechanism by which this occurs, the researchers found that in cultured oral cancer cells a protein called dynein is missing from the spindle, and the centrosomes no longer cluster together.
Furthermore, the researchers discovered that in some types of tumors, dynein is inhibited by the overexpression of another protein called NuMA. Excess NuMA seems to prevent dynein from binding to the spindle. When they reduced the level of NuMA in cultured cancer cells, the dynein returned to the spindles, and the spindles were no longer multipolar.
"This finding suggests that a possible treatment for some types of cancer could be a drug that inhibits NuMA," noted coauthor Susanne M. Gollin, professor of human genetics in Pitt's Graduate School of Public Health and coinvestigator at the Oral Cancer Center of Discovery at the University of Pittsburgh Cancer Institute.
On the Dynein conection:
The centrosome is a tiny cellular organ that helps organize chromosomes during cell division. It's also a signaling center for the cell, and as such is involved in the natural programmed cell death called apoptosis. In yet another role, the centrosome is important in setting up the two poles of the cell during mitosis, when cells segregate their chromosomal material and divide. Both these functions, when disrupted, can cause disease.
Dynamin-2 is an enzyme that helps cells form vesicles used to internalize agents from the cell's external environment. Through a series of laboratory manipulations of mammalian cells that included disrupting the centrosome, the research team was able to demonstrate that Dynamin-2 is a normal and necessary component of the centrosome. "When we reduced the levels of dynamin in cells, this led to an impairment of the centrosome," Dr. McNiven explains. The researchers also identified the part of Dynamin-2 that connects it to the centrosome. On the centrosome, they identified the part that recruits and binds Dynamin-2. "As a result of all this investigation, we now know that Dynamin-2 is vital to normal cell structure and organization," Dr. McNiven says.
More on abnormal spindles and centrosomes:
During cell division, spindle fibres align the 46 chromosomes along the middle of the cell and then forces are exerted that pull the chromosomes to opposite directions. A bi-polar spindle ensures that each of the two daughter cells receives a copy of each of the 46 chromosomes. Dr. Chatzimeletiou's team found that some spindles in cleavage and blastocyst stage embryos contain more than two poles (tripolar or tetraplolar), and in those it is impossible for the 46 chromosomes to be segregated equally to two daughter cells, as they would be pulled to three or more directions. "This in turn inevitably leads to chromosomal chaos", she saidGranted, this is more concered with fertility than cancer, but it does provide supporting evidence for the idea mentioned in Abnormal Spindles (above)
On the role of DNA Methylation:
This type of gene defect involves a process known as DNA methylation. Normally, healthy cells use DNA methylation during development and throughout life to turn off genes that are not needed. In cancer cells, aberrant DNA methylation turns off genes that would normally prevent inappropriate cell division. In other words, it shuts down one of the body's best mechanisms that prevents a cell from becoming cancerous.
In reality, cancer has a wide variety of causes and a lot of creative and interesting approaches are being taken in the study of cancer - none of them, however, involve Wells' "shaken centriole syndrome". Metaphors and analogies - such as comparing cells to machines may have a usefule heuristic purpose but if you take the comparison too seriously and assume cells are machines you usually end up looking silly.







